Introducción
En los últimos años, los inmunoestimulantes han ido reemplazando a los antibióticos por los impactos y problemas que generan en la producción de peces (Vásquez-Piñeros et al., 2012). Los inmunoestimulantes de origen natural incrementan la resistencia a enfermedades por la activación de la primera línea de defensa (inmune innata). Como los β-glucanos obtenidos de la pared celular de levaduras presentan actividades que mejoran la salud de peces (Miest et al., 2016). Para el β-glucano de la levadura Cystobasidium benthicum se ha descrito su posible uso en organismos como peces (Reyes-Becerril et al., 2021) e incluso humanos (Sanchez et al., 2021). Por lo tanto, este estudio evaluó la respuesta de defensa inmunes innatas generadas por el β-glucano de C. benthicum (βG-Cyben) en leucocitos del bazo de un pez emblemático del Golfo de California: la Totoaba macdonaldi.
Objetivo:
Evaluar la respuesta inmune generada por βG-Cyben en leucocitos del bazo de la totoaba.
Materiales y métodos
Obtención de βG-cyben: La levadura Cystobasidium benthicum LR192 se cultivó en el laboratorio para extraer el β-glucano usando: soluciones alcalinas (para separar la pared celular), soluciones ácidas (para eliminar grasas y proteínas), y etanol (para purificar el glucano), siguiendo la metodología descrita por Sánchez et al. (2021).
Aislamiento y estimulación de leucocitos: Los bazos de totoabas fueron donados por Earth Ocean Farms, La Paz, Baja California Sur, México. Los leucocitos se aislaron del bazo mediante técnicas estandarizadas (Reyes-Becerril et al., 2016); y se concentraron hasta obtener 1 millón de leucocitos por mililitro. Enseguida, se agregaron diferentes dosis de βG-cyben (50, 100 y 200 μg/mL de medio de cultivo celular) y se incubaron durante 24 horas a 25 °C (Machuca, 2021).
Análisis de respuestas de defensa: Primero, se analizó el número de células vivas (viabilidad) para verificar que βG-Cyben no causaba daño. Otros parámetros como la habilidad fagocítica, la actividad de explosión respiratoria, producción de óxido nítrico y la actividad de la enzima superóxido dismutasa (SOD) fueron medidos con técnicas estándar de laboratorio descritas y citadas por Machuca, (2021).
Resultados y discusión
El βG-Cyben tiene una proporción de 53.4 % de enlaces β-1,3–1,6-glucano y un peso molecular de 2.32 kDa. Se observó que las tres concentraciones de βG-Cyben no fueron tóxicas para los leucocitos del bazo de totoaba (Fig. 1) como reportaron Reyes-Becerril et al. (2021) en células del timo de T. macdonaldi.
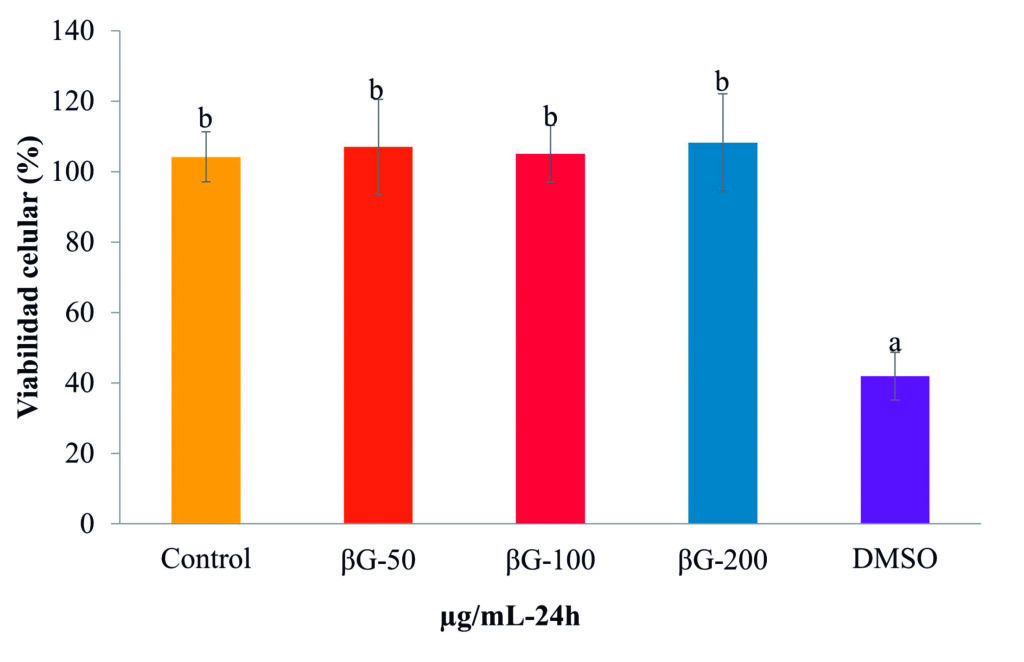
Figura 1. Viabilidad de leucocitos de bazo de Totoaba macdonaldi a las 24 h de estimulación con βG-cyben (50, 100 y 200 μg/mL). Las barras representan la media ± desviación estándar y las letras indican las diferencias significativas (p < 0.05) entre tratamientos. DMSO: Control de muerte celular (tóxico).
Los resultados indicaron que la concentración de 50 μg/mL de βG-Cyben (Fig. 2) es suficiente para activar la habilidad fagocítica, mecanismo de destrucción de patógenos por reacciones oxidativas realizado por los fagocitos (macrófagos y granulocitos). Este proceso y todas las actividades inmunes generadas por la estimulación de β-glucanos se origina en el reconocimiento de receptores transmembranales de tipo Toll y lectinas de tipo C (Pietretti et al., 2013). Este mismo efecto lo reportaron Callol et al. (2013) en leucocitos de la anguila europea (Anguilla anguilla) estimulados con 10 μg/mL del β-glucano comercial (Zymosan) obtenido de la levadura Saccharomyces cerevisiae. La producción del compuesto oxidativo y tóxico (óxido nítrico) para patógenos se presentó en leucocitos de bazo a medida que se aumentó la concentración de βG-cyben (Fig. 3).
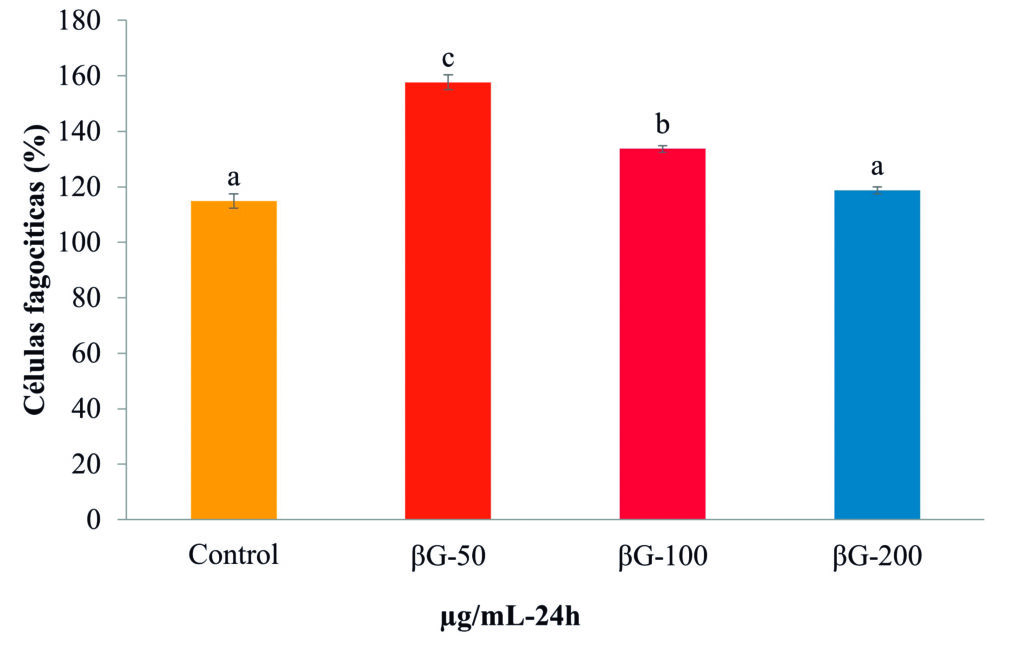
Figura 2. Células fagocíticas de bazo de Totoaba macdonaldi a las 24 h de estimulación con βG-cyben (50, 100 y 200 μg/mL). Las barras representan la media ± desviación estándar y las letras indican las diferencias significativas (p < 0.05) entre tratamientos.
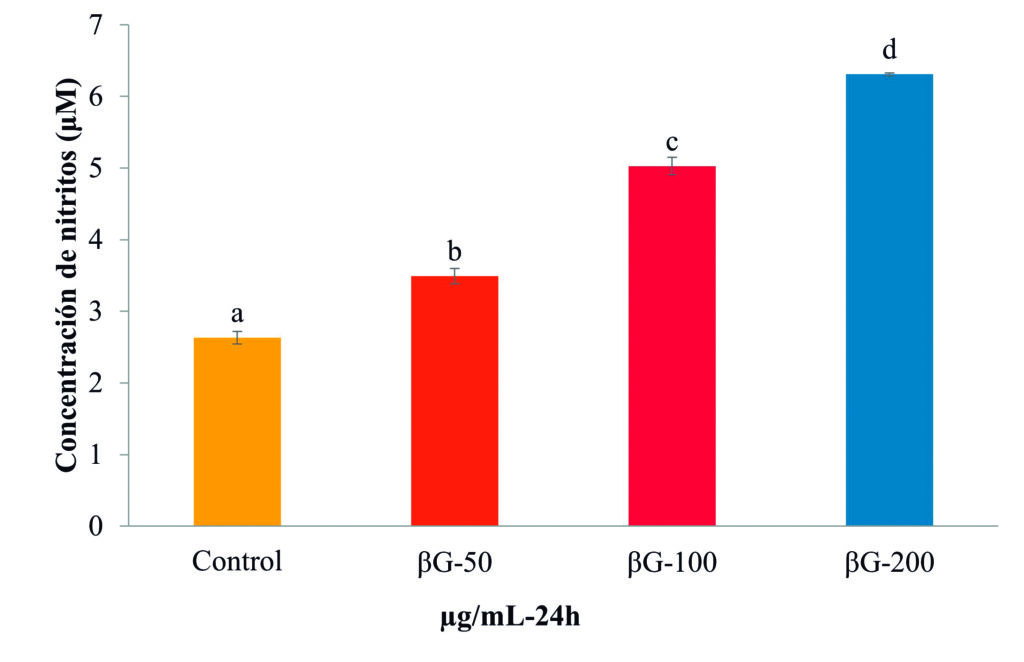
Figura 3. Producción de óxido nítrico (estimada por la concentración de nitritos) en leucocitos de bazo de Totoaba macdonaldi a las 24 h de estimulación con βG-cyben (50, 100 y 200 μg/mL). Las barras representan la media ± desviación estándar y las letras indican las diferencias significativas (p < 0.05) entre tratamientos.
Este efecto también se encontró en células del timo de totoaba estimuladas con βG-cyben (Reyes-Becerril et al., 2021). El proceso de generación de especies reactivas de oxígeno (eROS) para la producción de compuestos tóxicos, también conocido como explosión respiratoria se dio con la concentración de 200 μg/mL de βG-Cyben (Fig. 4).
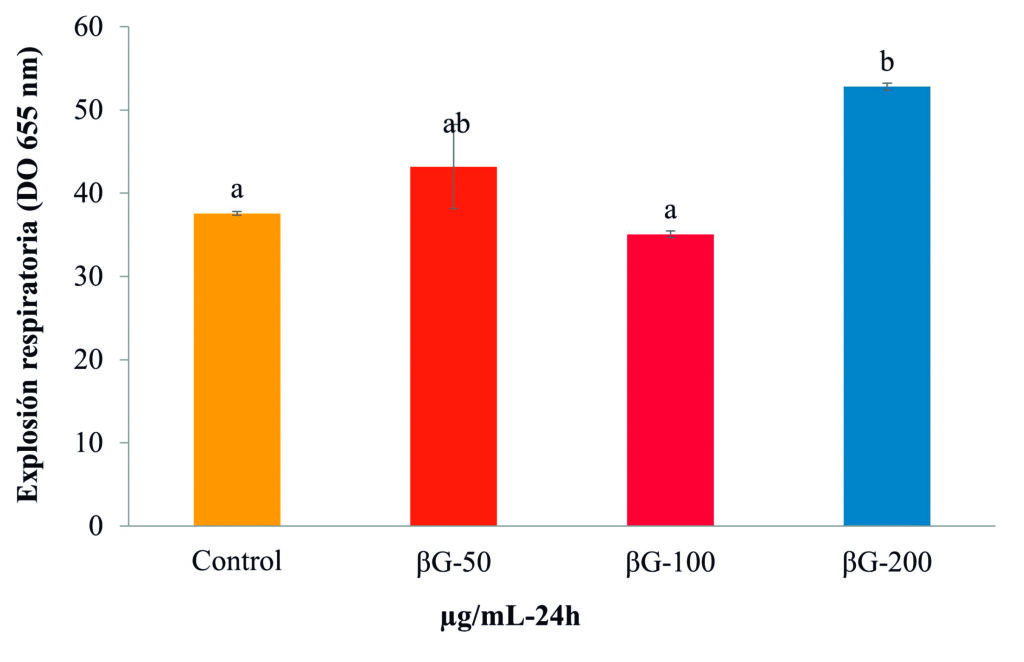
Figura 4. Actividad de explosión respiratoria en leucocitos de bazo de Totoaba macdonaldi a las 24 h de estimulación con βG-cyben (50, 100 y 200 μg/mL). Las barras representan la media ± desviación estándar y las letras indican las diferencias significativas (p < 0.05) entre tratamientos.
Este mismo efecto lo observó Vera-Jimenez et al. (2013) en leucocitos de la Carpa (Cyprinus carpio) con la concentración de 100 μg/mL de β-glucano comercial de S. cerevisiae (MacroGard). Como se producen especies tóxicas de oxígeno que pueden dañar las células del pez por estrés (Godahewa et al., 2015); entonces, para evitarlo, los leucocitos producen la enzima superóxido dismutasa (SOD) que tiene actividad antioxidante (Atli et al., 2016). En este trabajo, la actividad de SOD en leucocitos de totoaba aumentó a medida que se incrementó la concentración de βG-cyben (Fig. 5), sugiriendo una posible protección antioxidante.
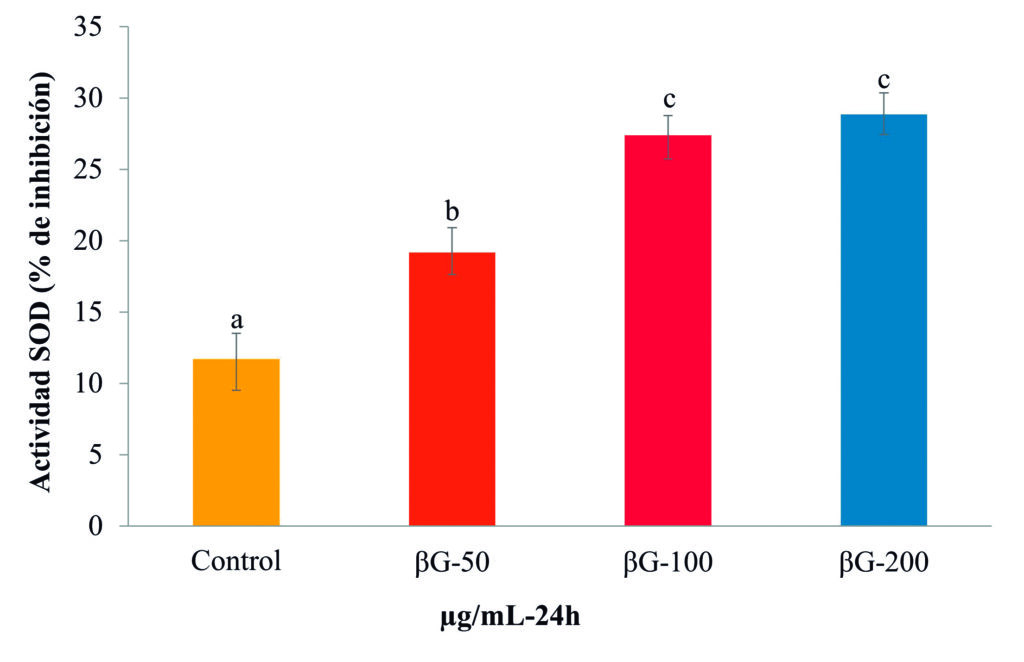
Figura 5. Actividad de superóxido dismutasa (SOD) en leucocitos de bazo de Totoaba macdonaldi a las 24 h de estimulación con βG-cyben (50, 100 y 200 μg/mL). Las barras representan la media ± desviación estándar y las letras indican las diferencias significativas (p < 0.05) entre tratamientos.
Conclusiones
El βG-cyben estimuló las actividades de defensa inmune analizadas en leucocitos, por lo que se sugiere su evaluación como inmunoestimulante para el cultivo de la totoaba.